Las briofitas, del latín bryophyta, derivado del griego βρύον bryon, musgo, y φυτόν phyton, planta, son plantas no vasculares y fueron las primeras en evolucionar hace 420 millones de años. Son descendientes de las algas verdes y fueron las primeras en colonizar los espacios terrestres, en esta división tenemos musgos, hepáticas y antóceras. Crecen en climas fríos o muy húmedos. En su cuerpo se distinguen 3 partes: rizoide, caulidios y filidios, análogos (pero no homólogos) a la raíz, tallo y hojas de los vegetales superiores. No poseen semillas.
Las briofitas con tejidos poco diferenciados no poseen vasos de conducción, es decir no tienen xilema ni floema, no presentan raíces, tallos ni hojas verdaderas, sino por un cuerpo vegetativo con estructuras muy primitivas, con células que no llegan a constituir un tejido. Así que las "raíces" que presentan se denominan rizoides, el "tallito", cauloide y las "hojitas", filoides. No está de más resaltar que debido al bajo nivel de organización que poseen estos organismos, ninguna de estas estructuras recién mencionadas son órganos verdaderos como sí aparecen en las traqueofitas o plantas vasculares.
En el ambiente terrestre, con cerca de 20,000 especies, las briofitas son el segundo grupo más importante de plantas verdes.
Generalmente son pequeñas y habitan en ambientes muy variados, desde cerca del nivel del mar hasta las elevaciones más altas, en las selvas o en los desiertos, pero su vida siempre está íntimamente ligada al agua en estado líquido. Su ciclo de vida incluye dos fases: el gametofito y el esporofito.
Cada una de ellas tiene atributos morfológicos y biológicos que señalan a las briofitas como un grupo excepcional y muy importante en la evolución del reino vegetal. Tradicionalmente se les divide en tres categorías, a saber, Antocerotes, Hepáticas y Musgos.
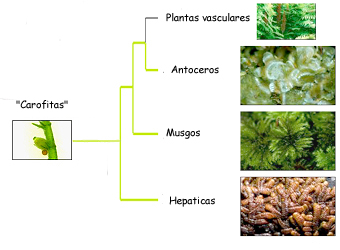
Esquema del árbol evolutivo de las plantas terrestres. Foto: University of Michiga, Neofronteras.
Clasificación tradicional
División BryophytaClase Anthocerotopsida - Antocerotes

Clase Hepaticopsida - Hepáticas

Clase Bryopsida - Musgos


Propuestas recientes
División Anthocerotophyta
División Marchantiophyta
Clase Marchantiopsida
Clase Jungermanniopsida
División Bryophyta
Clase Takakiopsida
Clase Sphagnopsida
Clase Andreaeopsida
Clase Andreaeobryopsida
Clase Polytrichopsida
Los estudios modernos indican que a pesar de estar relacionados, los grupos principales de briofitas son muy diferentes entre sí y como no tienen un origen común, deben ser clasificadas en divisiones independientes. El término `Briofita' tiene, en este sentido, una connotación informal; en las clasificaciones recientes, el término se refiere únicamente a los grupos de musgos.
Las briofitas son organismos muy antiguos de gran importancia científica. Se encuentran entre los primeros que ocuparon el ambiente terrestre; son clave en la evolución de las plantas terrestres, pero sus ligas con ellas son difíciles de establecer. El ciclo de vida de muchos artrópodos y microorganismos depende de los microambientes de las briofitas; muchas semillas de las plantas vasculares germinan en sus céspedes pues retienen agua y la liberan lentamente. Por esta característica también intervienen en el balance hídrico de los bosques y en la reducción de la erosión en ciertos ambientes. Su eliminación de bosques y selvas podría dar lugar a deterioro ecológico pues también parecen intervenir en el ciclo del carbono y otros minerales. Antes de calificarlos como insignificantes en la economía del hombre, se debe evaluar su papel ecológico y su utilidad como material de experimentación.
Las briofitas tienen un ciclo de vida heteromórfico. El gametofito o fase haploide es la dominante en las briofitas; es la más conspicua y la que tiene mayor duración. En comparación, el esporofito o fase diploide es pequeño y de duración corta. En la fase del gametofito, las plantas verdes pueden tener formas taloides o foliosas que derivan de la división de una sola célula apical. Durante la etapa reproductiva, el gametofito produce órganos sexuales masculinos (anteridios) o femeninos (arquegonios); las células sexuales masculinas o anterozoides son biflagelados y, por lo tanto, necesitan un ambiente acuoso para desplazarse. Los anterozoides y la oosfera (célula sexual femenina) se producen por mitosis pues se forman en gametofitos haploides. Al ocurrir la fecundación, la célula diploide o cigoto, derivada de la unión de las dos células sexuales, por divisiones sucesivas se transforma en un embrión multicelular. Más tarde, una vez diferenciado el pie que lo sujeta al gametofito y la seta o pedicelo que sostiene a una cápsula, el embrión se transforma en un esporofito en el que la división meiótica del tejido esporógeno de la cápsula permite la formación de esporas unicelulares haploides. Las esporas maduras son liberadas y al dispersarse, germinan y forman filamentos o masas celulares que se conocen como `protonema'. A partir del protonema se froman nuevos gametofitos.

Se conocen unas 100 especies de antocerotes en el mundo. Su gametofito es un talo multiestratificado en el que cada célula contiene de 1 a 12 cloroplastos usualmente lenticulares. En el centro del cloroplasto hay un pirenoide. La superficie inferior del talo tiene numerosos rizoides unicelulares y cuenta con poros que se comunican con cámaras de mucílago en las que se encuentran, con frecuencia, colonias de una alga del género Nostoc. Los anteridios y arquegonios se forman a partir de células superficiales dorsales; los anteridios están en cámaras mientras que la pared de los arquegonios es parte del tejido poco diferenciado del gametofito.

El esporofito de los antocerotes es una estructura cilíndrica con una pared de varias capas de células que encierran al tejido esporógeno y la columela. A partir del tejido esporógeno se forman las esporas y los pseudoelaterios; estos últimos son estructuras multicelulares intercaladas entre las esporas. La capa externa de la cápsula tiene estomas. Al madurar se forman de una a cuatro líneas de dehiscencia que se abren a partir del ápice y exponen a las esporas a la acción del viento. Un meristemo en la base de la cápsula permite el crecimiento y la producción de esporas indeterminados.
Los antocerotes parecen un grupo clave en la evolución de las plantas. El gametofito es de estructura simple y presenta rasgos primitivos como cloroplastos grandes y poco numerosos, con pirenoides (cuerpos proteicos que intervienen en la síntesis de almidón). En comparación, los estomas, la columela y el meristemo intercalar del esporofito pueden considerarse como rasgos avanzados. Los estomas aparecen aquí por primera vez e indican una adaptación al ambiente aéreo; la presencia de la columela indica el desarrollo progresivo de tejido estéril, una tendencia que se acentúa en otras plantas terrestres.
Se supone que existen entre 6500 a 7000 especies de hepáticas. Su gametofito puede ser taloide o folioso. El primero generalmente es postrado, como un listón, ramificado dicotómicamente, con una línea media y numerosos rizoides unicelulares en a superficie de contacto con el substrato. La complejidad anatómica de las formas taloides es variable, desde unas cuantas capas de células hasta las que muestran cámaras aéreas, poros, tejido fotosintético especial y tejidos de almacén. Los órganos sexuales generalmente se forman en la superficie superior del talo, con frecuencia sobre ramas especializadas. Los esporofitos, por tanto se desarrollan en las partes elevadas que favorecen la diseminación de las esporas por el viento. Los gametófitos pueden producir estructuras asexuales de reproducción como las yemas o propágulos que se forman en estructuras en forma de copa o media luna con características aerodinámicas peculiares. Las gotas de lluvia se encargan de liberarlas y dispersarlas a sitios cercanos.

El esporofito de las hepáticas tiene las tres partes fundamentales, pero es muy distinto al de los antocerotes. La pared carece de estomas y el tejido esporógeno forma elaterios unicelulares cuyos movimientos higroscópicos ayudan a la diseminación de las esporas. Las cápsulas se abren por valvas.
Las hepáticas son estructuralmente menos complejas que los musgos, pero en algunos lugares, especialmente en ciertos bosques tropicales, son más abundantes que los musgos. Algunas de ellas son frecuentes sobre las hojas de plantas vasculares. Una de sus peculiaridades fisiológicas es la producción de oleocuerpos que aparecen como pequeñas gotas en el citoplasma de las células.
Con cerca de 12,800 especies a nivel mundial, los musgos son el grupo más numeroso y el más diverso de las briofitas. El tallo puede ser erecto o postrado, de unos milímetros o centímetros de tamaño; su estructura interna, aunque simple, puede incluir un eje o cordón central en muchos musgos, y un sistema de conducción primitivo en algunos grupos. Además de los rizoides, los tallos de los musgos pueden tener estructuras fotosintéticas o de protección como pelos axilares, parafilios y pseudoparafilios. Las hojas están arregladas en forma espiral sobre el tallo, pero con frecuencia tienen otros arreglos; son uni o pluriestratificadas y sus células varían en tamaño, forma y ornamentación. Las hojas de muchos musgos tienen una costa, un nervio de longitud variable que puede prolongarse más allá de la lámina foliar en un mucrón o pelo foliar hialino. La costa puede se doble en algunos musgos tropicales y en algunos musgos, está cubierta por filamentos o lamelas. La anatomía de la costa también es compleja; consiste de varios tipos de células, incluyendo algunas de pared gruesa y otras de pared delgada que participan en la conducción de agua.
El esporofito de los musgos tiene las tres partes fundamentales, es decir, pie, seta y cápsula. El tamaño de la seta es variable y, en algunos grupos, está ausente. En Sphagnum es substituida por el pseudopodio, un pedicelo que se desarrolla a partir del tallo. La cápsula tiene una pared de varias capas de células y lleva estomas en la la parte basal o hipófisis. En el extremo distal se distingue la caliptra, una cubierta membranosa, fugaz, derivada del arquegonio. La boca de la cápsula está cubierta por una tapa u opérculo; al madurar, el opérculo cae y deja al descubierto 1 o 2 ciclos de apéndices que en conjunto son denominados `peristoma'. El peristoma regula la liberación de las esporas. El tejido esporógeno forma solamente esporas, pero la cápsula forma mayor cantidad de tejido estéril, incluyendo a la columela en un saco esporífero.

diferencias de las briofitas
![[tabla.bmp]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEi6pdtm8QU45Se5EB4ImLC5WpV4xfpxlGCJSbGE7F3ZNsX5Lry-CcRRJfLjppA6vf13_M1W1y_KDSJEDVRko9wTJ5u3-sTc2o6Whl63LKY9xYcY3uccQk_PeUIlAZtH4WYd02mP7Mn0FIvN/s1600/tabla.bmp)
pon la bibliografía
ResponderEliminar